 Old Web
Old Web
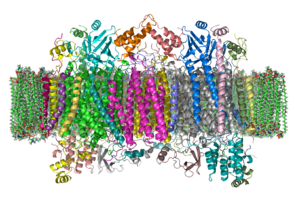
Cytochrome c oxidase

The enzyme cytochrome c oxidase or Complex IV, EC 1.9.3.1 is a large transmembrane protein complex found in bacteria, archaea, and in eukaryotes in their mitochondria.ETCComplex IV The enzyme cytochrome c oxidase or Complex IV, EC 1.9.3.1 is a large transmembrane protein complex found in bacteria, archaea, and in eukaryotes in their mitochondria. It is the last enzyme in the respiratory electron transport chain of cells located in the membrane. It receives an electron from each of four cytochrome c molecules, and transfers them to one dioxygen molecule, converting the molecular oxygen to two molecules of water. In this process it binds four protons from the inner aqueous phase to make two water molecules, and translocates another four protons across the membrane, increasing the transmembrane difference of proton electrochemical potential which the ATP synthase then uses to synthesize ATP. The complex is a large integral membrane protein composed of several metal prosthetic sites and 14 protein subunits in mammals. In mammals, eleven subunits are nuclear in origin, and three are synthesized in the mitochondria. The complex contains two hemes, a cytochrome a and cytochrome a3, and two copper centers, the CuA and CuB centers. In fact, the cytochrome a3 and CuB form a binuclear center that is the site of oxygen reduction. Cytochrome c, which is reduced by the preceding component of the respiratory chain (cytochrome bc1 complex, complex III), docks near the CuA binuclear center and passes an electron to it, being oxidized back to cytochrome c containing Fe3+. The reduced CuA binuclear center now passes an electron on to cytochrome a, which in turn passes an electron on to the cytochrome a3-CuB binuclear center. The two metal ions in this binuclear center are 4.5 Å apart and coordinate a hydroxide ion in the fully oxidized state. Crystallographic studies of cytochrome c oxidase show an unusual post-translational modification, linking C6 of Tyr(244) and the ε-N of His(240) (bovine enzyme numbering). It plays a vital role in enabling the cytochrome a3- CuB binuclear center to accept four electrons in reducing molecular oxygen to water. The mechanism of reduction was formerly thought to involve a peroxide intermediate, which was believed to lead to superoxide production. However, the currently accepted mechanism involves a rapid four-electron reduction involving immediate oxygen-oxygen bond cleavage, avoiding any intermediate likely to form superoxide.:865–866 COX assembly in yeast is a complex process that is not entirely understood due to the rapid and irreversible aggregation of hydrophobic subunits that form the holoenzyme complex, as well as aggregation of mutant subunits with exposed hydrophobic patches. COX subunits are encoded in both the nuclear and mitochondrial genomes. The three subunits that form the COX catalytic core are encoded in the mitochondrial genome. Hemes and cofactors are inserted into subunits I & II. The two heme molecules reside in subunit I, helping with transport to subunit II where two copper molecules aid with the continued transfer of electrons. Subunits I and IV initiate assembly. Different subunits may associate to form sub-complex intermediates that later bind to other subunits to form the COX complex. In post-assembly modifications, COX will form a homodimer. This is required for activity. Both dimers are connected by a cardiolipin molecule, which has been found to play a key role in stabilization of the holoenzyme complex. The dissociation of subunits VIIa and III in conjunction with the removal of cardiolipin results in total loss of enzyme activity. Subunits encoded in the nuclear genome are known to play a role in enzyme dimerization and stability. Mutations to these subunits eliminate COX function. Assembly is known to occur in at least three distinct rate-determining steps. The products of these steps have been found, though specific subunit compositions have not been determined. Synthesis and assembly of COX subunits I, II, and III are facilitated by translational activators, which interact with the 5’ untranslated regions of mitochondrial mRNA transcripts. Translational activators are encoded in the nucleus. They can operate through either direct or indirect interaction with other components of translation machinery, but exact molecular mechanisms are unclear due to difficulties associated with synthesizing translation machinery in-vitro. Though the interactions between subunits I, II, and III encoded within the mitochondrial genome make a lesser contribution to enzyme stability than interactions between bigenomic subunits, these subunits are more conserved, indicating potential unexplored roles for enzyme activity.
- CCF Conference Analysis
- Map Galaxy
- Academic Report
- What's New