 Old Web
Old Web
Ubiquitin ligase

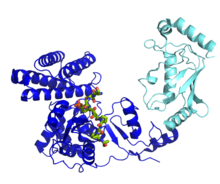
A ubiquitin ligase (also called an E3 ubiquitin ligase) is a protein that recruits an E2 ubiquitin-conjugating enzyme that has been loaded with ubiquitin, recognizes a protein substrate, and assists or directly catalyzes the transfer of ubiquitin from the E2 to the protein substrate. The ubiquitin is attached to a lysine on the target protein by an isopeptide bond. E3 ligases interact with both the target protein and the E2 enzyme, and so impart substrate specificity to the E2. Commonly, E3s polyubiquitinate their substrate with Lys48-linked chains of ubiquitin, targeting the substrate for destruction by the proteasome. However, many other types of linkages are possible and alter a protein's activity, interactions, or localization. Ubiquitination by E3 ligases regulates diverse areas such as cell trafficking, DNA repair, and signaling and is of profound importance in cell biology. E3 ligases are also key players in cell cycle control, mediating the degradation of cyclins, as well as cyclin dependent kinase inhibitor proteins. The human genome encodes over 600 putative E3 ligases, allowing for tremendous diversity in substrates. A ubiquitin ligase (also called an E3 ubiquitin ligase) is a protein that recruits an E2 ubiquitin-conjugating enzyme that has been loaded with ubiquitin, recognizes a protein substrate, and assists or directly catalyzes the transfer of ubiquitin from the E2 to the protein substrate. The ubiquitin is attached to a lysine on the target protein by an isopeptide bond. E3 ligases interact with both the target protein and the E2 enzyme, and so impart substrate specificity to the E2. Commonly, E3s polyubiquitinate their substrate with Lys48-linked chains of ubiquitin, targeting the substrate for destruction by the proteasome. However, many other types of linkages are possible and alter a protein's activity, interactions, or localization. Ubiquitination by E3 ligases regulates diverse areas such as cell trafficking, DNA repair, and signaling and is of profound importance in cell biology. E3 ligases are also key players in cell cycle control, mediating the degradation of cyclins, as well as cyclin dependent kinase inhibitor proteins. The human genome encodes over 600 putative E3 ligases, allowing for tremendous diversity in substrates. In enzymology, an ubiquitin-protein ligase (EC 6.3.2.19) is an enzyme that catalyzes the chemical reaction The 3 substrates of this enzyme are ATP, ubiquitin, and a lysine residue on a protein, whereas its 3 products are AMP, diphosphate, and protein N-ubiquityllysine. Canonical ubiquitylation creates an isopeptide bond between a lysine residue on a target protein and the ubiquitin C-terminal Glycine 76. This enzyme belongs to the family of ligases, to be specific those forming carbon-nitrogen bonds as acid-D-amino-acid ligases (peptide synthases). The systematic name of this enzyme class is ubiquitin:protein-lysine N-ligase (AMP-forming). This enzyme is also called ubiquitin-activating enzyme. This enzyme participates in 3 metabolic pathways: ubiquitin-mediated proteolysis, Parkinson's disease, and Huntington's disease. The ubiquitin ligase is referred to as an E3, and operates in conjunction with an E1 ubiquitin-activating enzyme and an E2 ubiquitin-conjugating enzyme. There is one major E1 enzyme, shared by all ubiquitin ligases, that uses ATP to activate ubiquitin for conjugation and transfers it to an E2 enzyme. The E2 enzyme interacts with a specific E3 partner and transfers the ubiquitin to the target protein. The E3, which may be a multi-protein complex, is, in general, responsible for targeting ubiquitination to specific substrate proteins. The ubiquitylation reaction proceeds in three or four steps depending on the mechanism of action of the E3 ubiquitin ligase. In the conserved first step, an E1 cysteine residue attacks the ATP-activated C-terminal glycine on ubiquitin, resulting in a thioester Ub-S-E1 complex. The energy from ATP and diphosphate hydrolysis drives the formation of this reactive thioester, and subsequent steps are thermoneutral. Next, a transthiolation reaction occurs, in which an E2 cysteine residue attacks and replaces the E1. HECT domain type E3 ligases will have one more transthiolation reaction to transfer the ubiquitin molecule onto the E3, whereas the much more common RING finger domain type ligases transfer ubiquitin directly from E2 to the substrate. The final step in the first ubiquitylation event is an attack from the target protein lysine amine group, which will remove the cysteine, and form a stable isopeptide bond. One notable exception to this is p21 protein, which appears to be ubiquitylated using its N-terminal amine, thus forming a peptide bond with ubiquitin. Humans have an estimated 500-1000 E3 ligases, which impart substrate specificity onto the E1 and E2. The E3 ligases are classified into four families: HECT, RING-finger, U-box, and PHD-finger. The RING-finger E3 ligases are the largest family and contain ligases such as the anaphase-promoting complex (APC) and the SCF complex (Skp1-Cullin-F-box protein complex). SCF complexes consist of four proteins: Rbx1, Cul1, Skp1, which are invariant among SCF complexes, and an F-box protein, which varies. Around 70 human F-box proteins have been identified. F-box proteins contain an F-box, which binds the rest of the SCF complex, and a substrate binding domain, which gives the E3 its substrate specificity. Ubiquitin signaling relies on the diversity of ubiquitin tags for the specificity of its message. A protein can be tagged with a single ubiquitin molecule (monoubiquitylation), or variety of different chains of ubiquitin molecules (polyubiquitylation). E3 ubiquitin ligases catalyze polyubiquitination events much in the same way as the single ubiquitylation mechanism, using instead a lysine residue from a ubiquitin molecule currently attached to substrate protein to attack the C-terminus of a new ubiquitin molecule. For example, a common 4-ubiquitin tag, linked through the lysine at position 48 (K48) recruits the tagged protein to the proteasome, and subsequent degradation. However, all seven of the ubiquitin lysine residues (K6, K11, K27, K29, K48, and K63), as well as the N-terminal methionine are used in chains in vivo. Monoubiquitination has been linked to membrane protein endocytosis pathways. For example, phosphorylation of the Tyrosine at position 1045 in the Epidermal Growth Factor Receptor (EGFR) can recruit the RING type E3 ligase c-Cbl, via an SH2 domain. C-Cbl monoubiquitylates EGFR, signaling for its internalization and trafficking to the lysosome.
- CCF Conference Analysis
- Map Galaxy
- Academic Report
- What's New